
Analysis of Growth and Leaf Morphological Variation of Ormosia hosiei from Different Origins and Selection of Excellent Strains
-
摘要:
揭示不同产地红豆树Ormosia hosiei家系生长与叶形的表型分化程度与变异规律,为筛选优良家系提供理论指导。本研究选择来自江西、浙江、福建和四川4省共38个红豆树家系,通过表型测定开展生长和叶形性状的差异分析和多样性研究。结果表明,不同产地红豆树生长性状与叶形性状存在极显著差异(P<0.01),所有表型的变异系数均超过10%,说明各表型性状在产地间均有丰富的变异。其中四川产地的红豆树表型特征明显,生长较差,其树高(3.01 m)、东西冠幅(1.31 m)和南北冠幅(1.35 m)均显著低于其他三个省份产地的红豆树(P<0.01),但叶片发育较好,叶面积(23.56 cm2)、叶周长(16.21 cm)、叶长(11.57 cm)和叶宽(5.50 cm)等表型均显著高于其他三个产地的红豆树(P<0.01),说明四川产地的红豆树家系整体长势一般,但叶片较大;表型间相关性分析结果显示生长性状和叶形性状内两两之间均存在极显著正相关(P<0.01),这为优异种质定向筛选提供依据。聚类分析结果将红豆树群体分为5类,聚类结果与地理位置基本一致。最终基于树高、胸径和冠幅表型数据筛选得到优良红豆树家系8个。
Abstract:By revealing the extent and pattern of phenotypic differentiation in growth and leaf morphology among different provenances of O. hosiei, this study provides theoretical guidance for selecting superior genetic resources. Thirty-eight O. hosiei provenances from four provinces (Jiangxi, Zhejiang, Fujian, and Sichuan) were selected for phenotypic characterization of growth and leaf traits through differential analysis and diversity study. Results showed significant variations in growth and leaf traits among different provenances, with all phenotypic coefficients of variation exceeding 10%, indicating rich variability across provenances for all traits. Specifically, O. hosiei from Sichuan exhibited distinctive phenotypic characteristics, with inferior growth traits including tree height (3.01 m), east-west canopy width (1.31 m), and north-south canopy width (1.35 m) significantly lower than those from the other three provinces (p<0.01), but superior leaf development traits including leaf area (23.56 cm2), leaf perimeter (16.21 cm), leaf length (11.57 cm), and leaf width (5.50 cm) significantly higher than those from the other three provenances (p<0.01), indicating moderate overall growth but larger leaves for Sichuan provenances. Correlation analysis revealed highly significant positive correlations between growth and leaf traits, providing a basis for targeted selection of superior germplasm. Cluster analysis categorized O. hosiei populations into five groups, which largely corresponded to their geographical origins. Finally, eight superior O. hosiei provenances were identified based on growth trait data including tree height, diameter at breast height, and canopy width, providing theoretical and material foundations for the breeding of elite O. hosiei varieties.
-
Keywords:
- O. hosiei /
- growth traits /
- leaf morphological traits /
- variance portion /
- cluster analysis
-
红豆树Ormosia hosiei为豆科Leguminosae红豆属Ormosia常绿或半落叶乔木,是我国特有的珍贵阔叶用材树种,自然分布于浙江、江西、福建、四川、陕西等省[1-2]。红豆树木材坚实硬重,结构细密,不经油漆却形如墨玉,质地优良,具有极高的材用、景观和森林文化价值[3-4],闻名于世的龙泉宝剑剑鞘就是用其心材加工而成。目前,对红豆树的研究主要集中在生物学特性[5]、遗传分布[1]、育苗造林[6]、人工林经营[7]、分子标记[8]等方面。考虑到气候条件、人工定向选择等因素的影响,目前对多个省份产地的红豆树优良家系的选择还少有研究。
遗传评价与选择是林木育种的重要方法[9],其评价结果可为选育地区提供可靠的应用材料,对于实现适地适种具有重要应用意义[10-11]。生长性状和叶形性状作为林木中最直观的表型性状,受到环境因素和遗传因素共同作用,是林木家系评价与选择的关键因素[12],表型性状在家系间存在丰富的变异[13-14]。陈清根[15]开展了不同种源红豆树遗传变异的研究,通过对8年生林龄7个不同种源的红豆树树高、胸径和单株材积等生长性状进行测定,在闽南地区开展种源试验,筛选出适合当地栽植的优良红豆树种源。崔珺等[16]对9个不同地理种源的红豆树幼苗叶片功能性状和碳氮磷含量进行分析测定,对红豆树幼苗的地理环境适应性提出见解,最终确定在当地引种区适合的种源材料。李峰卿等[17]对4个省份共76份红豆树优树种子和幼苗性状进行了测定并初步筛选优良家系。但利用优良家系、单株评价开展后向选择研究,扩大红豆树优良家系的选择范围,可以更有效地揭示林木主要性状的遗传变异规律并为科学遗传改良提供理论依据。因此,本研究通过收集不同产地、不同家系的红豆树家系苗,分析测定其表型性状以筛选表型性状优异的家系,以保证红豆树的有效利用。
本研究以4个产地38个家系的红豆树为研究材料,重点开展对不同产地、不同家系红豆树的生长情况、分枝、冠幅、叶形等表型性状的遗传变异规律的研究,建立生长性状与叶形性状的相关性,揭示不同产地、不同家系红豆树生长性状与叶形性状的关系与差异情况,选育一批优质高产的红豆树材料,为红豆树优良家系的繁育提供理论和材料支撑。
1. 材料与方法
1.1 试验材料
本研究共收集红豆树家系苗38份,其中5份家系苗来源于江西省,15份家系苗来源于浙江省,15份家系苗来源于福建省,3份家系苗来源于四川省,具体材料来源见表1。所有家系苗均为种子萌发的实生苗,于2016年培育2年生容器苗,2017年培育1年生容器苗,并于2018年春定植于浙江省龙泉市林业科学研究院上圩基地。试验林随机区组设计,3次重复,株行距2.0 m×2.0 m,定植期间的前三年进行抚育管理,秋季施复合肥一次。
表 1 红豆树采种产地与种质来源Table 1. Originsites and seed groups of O. hosiei编号 来源地 采集地点 编号 来源地 采集地点 JX1 江西省 抚州1号马头山镇 FJ1 福建省 南平市茫荡山自然保护区 JX2 江西省 抚州2号马头山镇 FJ2 福建省 南平市浦城县石陂镇 JX3 江西省 抚州zx6马头山镇 FJ3 福建省 南平市浦城县临江镇 JX4 江西省 抚州4号马头山镇 FJ4 福建省 宁德市柘荣县富溪镇 JX5 江西省 抚州尖峰1号广昌尖峰乡 FJ5 福建省 宁德市柘荣县宅中乡 ZJ1 浙江省 龙泉市锦溪镇 FJ6 福建省 宁德市柘荣县东源乡 ZJ2 浙江省 龙泉市八都镇 FJ7 福建省 宁德市柘荣县富溪镇 ZJ3 浙江省 龙泉市林业科学研究院 FJ8 福建省 宁德市柘荣县宅中乡 ZJ4 浙江省 龙泉市林业科学研究院 FJ9 福建省 宁德市福安市下白石镇 ZJ5 浙江省 龙泉市林业科学研究院 FJ10 福建省 宁德市福安市下白石镇 ZJ6 浙江省 龙泉市林业科学研究院 FJ11 福建省 宁德市柘荣县富溪镇 ZJ7 浙江省 龙泉市外窟 FJ12 福建省 宁德市柘荣县富溪镇 ZJ8 浙江省 龙泉市林业局 FJ13 福建省 宁德市柘荣县富溪镇 ZJ9 浙江省 龙泉市杨梅尖 FJ14 福建省 宁德市柘荣县富溪镇 ZJ10 浙江省 龙泉市八都镇 FJ15 福建省 宁德市柘荣县富溪镇 ZJ11 浙江省 龙泉市剑池街道 SC1 四川省 资阳市安岳县李家镇 ZJ12 浙江省 龙泉市江滨路 SC2 四川省 内讧市太平镇 ZJ13 浙江省 丽水市云和县西弄村 SC3 四川省 内讧市太平镇 ZJ14 浙江省 丽水市庆元县黄田镇 ZJ15 浙江省 丽水市庆元县黄田镇 1.2 试验地点
本试验在浙江省龙泉市林科院上圩基地开展,上圩基地属亚热带季风气候,地理坐标为119°07'E、28°04'N。年平均气温17.6 ℃,年降水量1 699.4 mm,无霜期263 d,全年日照时数1849.8 h,立地条件较优,土壤肥力中等偏上。
1.3 试验方法
于2022年10月开展树高、胸径、冠幅和分枝数调查,同时,每个在重复的每个家系随机采集4个单株的叶片,每株树采集树冠中部10张完全功能叶片,放入封口袋中带回实验室,使用扫描仪(日本,Epson Expression 11000XL)测定叶周长、叶长、叶宽和叶面积等叶形性状。
1.4 数据分析
利用Excel 2016软件进行数据记录与基本统计,使用SPSS 25.0软件进行差异性分析和相关性分析,使用SPSS 25.0软件中的Ward法基于生长和叶形性状对红豆树家系进行聚类分析,使用R语言对数据进行双因素巢式分组设计方差分析和聚类分析。
采用双因素巢式设计方差分析,线性模型为:
$$ \mathit{Y} _{ \mathit{ijk} } =\mu + \mathit{S} _{ \mathit{i} } \mathrm+ \mathit{T} _{ \mathrm{(} \mathit{i} \mathrm{)} \mathit{j} } \mathrm{+\varepsilon (} \mathit{ij} \mathrm{)} \mathit{k} $$ (1) 式中,Yijk为第i个群体第j个家系第k个观测值,μ为总均值,Si为群体效应(固定),T(i)j为群体内家系效应(随机),ε (ij)k为试验误差[17]。
$$ \mathit{V} _{ \mathit{ST} } =\mathrm{(} \mathit{\sigma } ^{ \mathrm{2}} \mathit{t} / \mathit{s} \mathrm{)/(} \mathit{\sigma } ^{ \mathrm{2}} \mathit{t} \mathrm{/} \mathit{s} \mathrm+ \mathit{\sigma } ^{ \mathrm{2}} \mathit{s} ) $$ (2) 式中,VST为表型分化系数,表示群体间变异占遗传总变异的百分比[12-13],σ2t/s为群体间方差分量, σ2s为群体内方差分量[18-19]。
性状变异系数 (CV) 表示性状离散程度[18-20],式中Mean为性状平均值,SD为标准差。
$$ \mathit{CV} \mathrm{(\text{%})} =\mathit{SD} \mathrm{/} \mathit{Mean} \mathrm{*100\text{%} } $$ (3) 采用DPS数据处理系统9.01软件中的TOPSIS分析法综合评价并筛选红豆树优良单株。
2. 结果与分析
2.1 红豆树家系生长性状和叶形性状的产地遗传变异
通过对38份红豆树家系的树高、胸径、冠幅、叶面积、叶形等9个表型性状进行测定(表2),发现各表型性状在不同产地间均存在极显著差异(P<0.01)。来自四川的红豆树家系树高、胸径、东西冠幅、南北冠幅和分枝数量等生长性状分别为3.01 m、3.07 cm、1.31 m、1.35 m和7.06个,均极显著小于江西、浙江和福建产地的红豆树(P<0.01),但叶面积(23.56 cm2)、叶周长(16.21 cm)、叶长(11.57 cm)和叶宽(5.50 cm)等叶形性状均极显著高于江西、浙江和福建产地的表型性状(P<0.01)。这说明四川产地的红豆树家系整体长势一般但叶片生长较好,而浙江产地红豆树家系则表现出相反的规律,即长势良好但叶形较小。
表 2 不同产地红豆树生长性状和叶形性状的测定Table 2. Determination of growth traits and leaf morphological traits in different O. hosiei populations表型性状 江西 浙江 福建 四川 树高/m 3.36±0.09b 4.03±0.11a 4.12±0.05a 3.01±0.13b 胸径/cm 3.60±0.13b 4.16±0.07a 4.16±0.09a 3.07±0.15c 东西冠幅/m 1.76±0.06b 1.89±0.03ab 1.95±0.03a 1.31±0.07c 南北冠幅/m 1.72±0.06a 1.89±0.03a 1.91±0.03a 1.35±0.09b 分枝数量/个 7.51±0.26ab 8.20±0.16a 8.26±0.13a 7.06±0.30b 叶面积/cm2 15.55±0.39b 15.42±0.20b 16.78±0.25b 23.56±0.76a 叶周长/cm 13.33±0.18bc 13.09±0.11c 13.91±0.12b 16.21±0.30a 叶长/cm 9.65±0.09bc 9.37±0.05c 9.83±0.06b 11.57±0.19a 叶宽/cm 4.05±0.05c 4.19±0.03bc 4.28±0.03b 5.50±0.08a 注:数据列中不同小写字母表示在P<0.01水平存在极显著差异。 采用双因素巢式分组设计对产地间和家系间的表型性状进行方差分析,探究红豆树生长性状与叶形性状的差异性(表3)。由表3可知,除冠幅性状(Fp=0.864,Fi=1.096)外,其他生长性状和叶形性状在产地间和家系间均存在极显著差异(P<0.01),且产地间的F值均大于家系间的F值(Fp>Fi),说明除冠幅外的其他性状均在产地间和家系间存在丰富的多样性和广泛变异,具有较强的可选择性。
表 3 红豆树生长性状和叶形性状在产地间和家系间的方差分析Table 3. Analysis of variance on growth and leaf morphological traits among O. hosiei populations and individuals性状 产地间F值(Fp) 家系间F值(Fi) 树高 18.902** 2.774** 胸径 24.674** 11.881** 冠幅 0.864 1.096 分枝数 6.707** 3.712** 叶面积 52.184** 10.126** 叶周长 33.842** 7.118** 叶长 28.028** 13.881** 叶宽 106.173** 19.567** 注: **表示产地间和家系间均存在极显著差异(P<0.01)。 2.2 红豆树家系生长性状和叶形性状的变异
表4呈现了红豆树生长性状和叶形性状在不同产地间的变异情况,测定的9个性状均表现出不同程度的变异,各产地内红豆树表型性状的变异系数均大于10%,说明生长性状和叶形性状在不同产地间均表现出丰富的变异。其中叶长的变异程度最小(11.63%~15.68%),南北冠幅的变异程度最大( 25.93%~56.90%)。通过比对表型性状的变异系数发现,四川产地的红豆树生长性状和叶形性状均表现出更大的变异系数,其中南北冠幅和东西冠幅的变异系数最大,分别为56.90%和47.82%;叶周长、叶长和叶宽的变异系数相对较小,分别为17.28%、15.68%和13.95%。而福建产地的红豆树生长性状和叶形性状整体变异系数相对较小,变异系数最大的表型性状为胸径,为38.90%;变异系数最小的表型性状同样为叶周长、叶长和叶宽,分别为18.31%、13.41%和15.64%。这说明四川产地红豆树南北冠幅和东西冠幅表现出更显著的差异与更大的变异幅度,叶周长、叶长和叶宽在不同产地中均表现出较低的变异系数。
表 4 红豆树4个产地生长性状与叶形性状变异系数Table 4. Coefficient of variation of growth and leaf morphological traits among O. hosiei populations来源地 树高/% 胸径/% 东西冠幅/% 南北冠幅/% 分枝数/% 叶面积/% 叶周长/% 叶长/% 叶宽/% 江西 28.51 37.85 34.85 34.15 36.03 30.57 16.86 11.67 15.22 浙江 26.80 33.14 33.47 33.72 35.59 27.62 17.59 11.63 13.67 福建 22.94 38.90 27.64 25.93 30.64 31.24 18.31 13.41 15.65 四川 36.02 41.43 47.82 56.90 35.94 30.78 17.28 15.68 13.95 2.3 红豆树家系苗产地间和产地内表型性状分化
方差分量的占比和表型分化系数结果如表5所示。其中,树高、冠幅、叶面积、叶周长、叶长和叶宽等性状的产地间方差分量显著大于产地内方差分量,几种性状的表型分化系数变幅在70.89%~84.23%。而胸径和分枝数两个表型的方差分量在产地内更大,其表型分化系数分别仅为9.10%和12.41%,其中胸径性状的方差分量在产地内占比29.59%,产地内变异是胸径表型变异的主要来源,而大部分性状的表型变异主要来源于产地间,产地间的多样性大于产地内的多样性。
表 5 红豆树表型性状产地间和产地内的方差分量与表型分化系数Table 5. Variance portion and phenotypic differentiation coefficient (VST) among population性状 方差分量 方差分量百分比/% 表型分化系数(VST)/% 产地间(σ2t/s) 产地内(σ2s) 产地间(σ2t/s) 产地内(σ2s) 树高 0.24 0.06 19.22 5.04 79.21 胸径 0.07 0.68 2.96 29.59 9.10 东西冠幅 0.07 0.02 16.86 5.62 75.00 南北冠幅 0.06 0.02 13.79 5.67 70.89 分枝数 0.10 0.73 1.38 9.73 12.41 叶面积 13.68 2.56 35.02 6.56 84.23 叶周长 1.79 0.48 22.96 6.17 78.81 叶长 0.88 0.27 35.30 10.82 76.55 叶宽 0.41 0.08 49.88 9.98 83.33 2.4 红豆树生长性状与叶形性状的相关性分析
通过对9个红豆树表型性状的相关性进行分析(表6),发现生长性状树高、胸径、冠幅和分枝数两两之间均存在极显著正相关(P<0.01,r=0.171~0.783),4个叶形性状(叶面积、叶周长、叶长和叶宽)两两之间也存在极显著正相关(P<0.01,r=0.460~0.879)。生长性状与叶形性状之间的相关性分析结果发现,胸径与叶面积、叶周长、叶长和叶宽均存在负相关,r值分别为−0.042、−0.001、−0.004和−0.078,其中仅与叶宽呈显著负相关(P<0.05)。此外,树高与叶周长呈显著正相关(P<0.05),叶宽也与分枝数呈负相关,但并不显著。
表 6 红豆树表型性状相关性分析Table 6. The correlation between traits in the O. hosiei populations树高 胸径 东西冠幅 南北冠幅 分枝数 叶面积 叶周长 叶长 叶宽 树高 1.000 胸径 0.477** 1.000 东西冠幅 0.547** 0.681** 1.000 南北冠幅 0.491** 0.617** 0.783** 1.000 分枝数 0.171** 0.252** 0.317** 0.303** 1.000 叶面积 0.039 −0.042 0.019 0.013 0.014 1.000 叶周长 0.071* −0.001 0.026 0.016 0.028 0.879** 1.000 叶长 0.042 −0.004 0.028 0.030 0.006 0.667** 0.634** 1.000 叶宽 0.033 −0.078* 0.018 0.034 −0.025 0.724** 0.460** 0.622** 1.000 注:**表示极显著相关(P<0.01),*表示显著相关(P<0.05)。 2.5 红豆树生长性状和叶形性状聚类分析
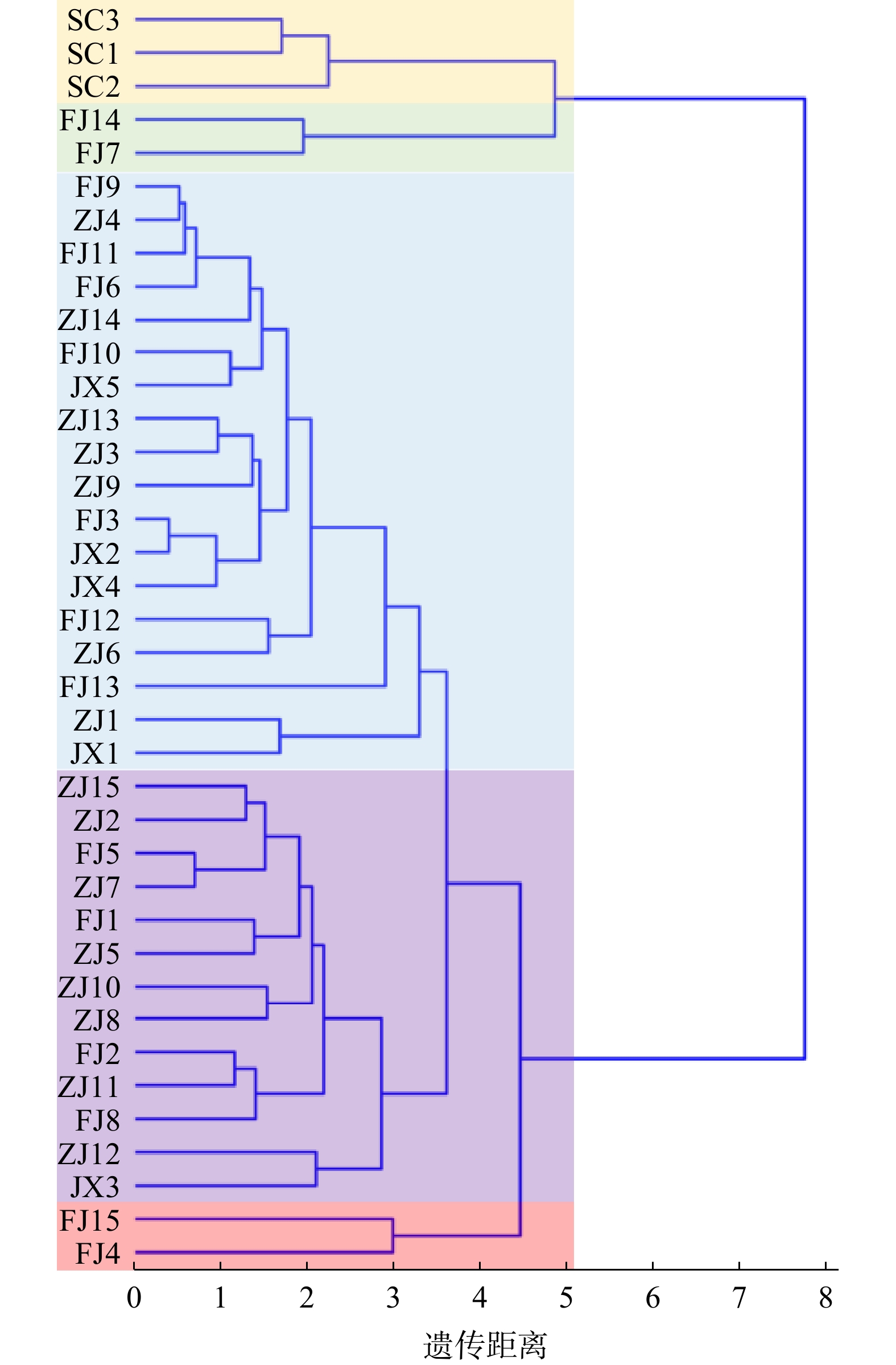
聚类分析结果表明,38个红豆树家系被分为5大类,其中江西、浙江和福建产地大部分的红豆树家系聚在一起,以SC3、SC1和SC2为代表的3个四川家系单独聚为一类(图1)。此外,FJ14和FJ7,FJ15和FJ4分别聚为一类,且与福建产地大多数的红豆树家系聚类结果不同,说明福建产地的红豆树家系差异相对较大。
![]() 图 1 基于9种表型的红豆树不同家系表型树状聚类图SC-四川;FJ-福建;ZJ-浙江;JX-江西。Figure 1. Cluster analysis of different populations of O. hosiei based on 9 phenotypic traits
图 1 基于9种表型的红豆树不同家系表型树状聚类图SC-四川;FJ-福建;ZJ-浙江;JX-江西。Figure 1. Cluster analysis of different populations of O. hosiei based on 9 phenotypic traits2.6 红豆树优良家系筛选
根据上述分析结果,选择胸径、树高和冠幅3个性状作为表型指标用于红豆树优良家系的筛选研究。采用TOPSIS法对家系进行综合排序,其中胸径和树高的权重值为2,冠幅的权重值为1。优良家系选择按照20%入选率进行,最终选出ZJ6、FJ14、FJ12、ZJ5、ZJ8、FJ13、FJ8和FJ6共8个优良家系(表7)。
表 7 8个优良家系的表型指标及排名Table 7. Phenotypic indicators and rankings of 8 excellent families of O. hosiei家系号 树高/m 胸径/cm 东西冠幅/m 南北冠幅/m 指标(CI) 排名 ZJ6 4.44±0.19a 5.23±0.29a 2.36±0.10a 2.38±0.11a 0.9564 1 FJ14 4.63±0.20a 5.26±0.24a 2.19±0.11a 2.09±0.10ab 0.9224 2 FJ12 4.56±0.18a 5.14±0.26a 2.13±0.10a 2.09±0.07ab 0.9080 3 ZJ5 4.42±0.16a 4.87±0.22a 2.21±0.10a 2.19±0.09ab 0.8853 4 ZJ8 4.23±0.19a 4.93±0.23a 2.13±0.10a 2.18±0.10ab 0.8629 5 FJ13 4.38±0.23a 4.97±0.32a 2.19±0.14a 1.95±0.14b 0.8621 6 FJ8 4.28±0.18a 4.99±0.28a 1.99±0.11a 2.02±0.10ab 0.8447 7 FJ6 4.09±0.13a 4.74±0.20a 1.94±0.08a 1.97±0.07ab 0.7841 8 入选均值 4.38 5.02 2.14 2.11 注:同一列中不同小写字母表示在P<0.01水平存在极显著差异。 这8个优良家系的树高和胸径分别为4.23~4.63 m和4.87~5.26 cm,且家系间均不存在显著差异。冠幅为1.94~2.38 m,其中仅ZJ6和FJ13的南北冠幅之间存在极显著差异(P<0.01)。
3. 讨论与结论
3.1 红豆树生长和叶形性状的遗传变异
4个产地红豆树家系之间的生长性状和叶形性状存在极显著差异(P<0.01),表明优良家系的选择具有良好的基础,可以为红豆树的生长性状和叶形性状提供更多的可能性。四川产地的红豆树家系总体生长表现一般,但叶片生长表现较江西、浙江和福建产地的红豆树家系更优(表2、表3)。表型间的相关性分析结果显示生长性状和叶形性状各自内部两两之间均存在极显著正相关(P<0.01),但胸径与叶形均存在负相关(表6),表明红豆树的叶片生长会导致树体内营养物质分布不均,影响植株胸径的发育进而影响植株的生长。
遗传变异是选择的基础,充分了解林木的变异才能准确地筛选优良种质,同时变异系数的大小也可以作为性状选择改良潜力的一个重要指标,决定着种质资源选择余地的大小[21]。长期的自然选择因生长环境的差异而产生家系和产地的互作。穆怀志等[22]发现糠椴 Tilia mandshurica天然居群的叶片和果实形状在居群间和居群内均存在极显著差异,且变异系数介于20.0%~39.9%。程琳等[23]发现杉木木材品质性状的变异系数为8.69%~40.95%,同样具有丰富的变异。在本研究中,4个产地红豆树的表型性状变异系数均较大(表4),变幅为11.63%~56.90%,说明红豆树在生长性状和叶形性状上均存在较大的改良潜力,有利于优良家系的评价和筛选[24]。此外,本研究红豆树中仅胸径和分枝数两种性状的变异主要源自产地内,而其他表型性状的变异主要源自产地间(表5)。生物群体间的变异反映了其对不同生存环境的适应程度,值越大适应的生态环境越广[25]。研究中选取的4个不同省份产地的红豆树,产地间的变异一定程度上反映了地理隔离层面上的变异,是种内多样性的重要组成部分[26]。
聚类分析结果显示,江西、浙江和福建产地的大部分红豆树家系聚在一起,根据地理分布及我国红豆树群体的划分,江西、浙江与福建三省在地理位置上相互毗邻,因此其生长和叶形性状等表型更相近,表型聚类聚在一起的结果是比较可靠的,说明地理位置相近的群体亲缘关系也较近,体现了数量性状变异的连续性。此外,聚类结果也发现部分福建产地的家系与四川产地的家系亲缘关系较近,这可能是因为种苗调拨或异地引种繁殖等因素的影响[16]。对不同地理位置蒙古栎Quercus mongolica 和板栗Castanea mollissima的遗传变异研究中发现相似的结果,王娜等[27]对蒙古栎种群多样性研究中发现6个蒙古栎种群按省份聚成了三类,刘亚斌等[28]对6个省份板栗刺苞和坚果形态多样性进行了分析,发现聚类结果与地理分布格局基本吻合。
3.2 红豆树优良家系的选择
相关性分析可以更清晰地明确不同性状间的关联程度[29],林木中大部分性状均为数量性状,其中生长性状和叶形性状间的相关性分析对红豆树种质资源的遗传改良具有重要意义。本研究中的生长和叶形两类性状内部两两之间均呈现极显著的正相关,但两类表型性状之间无显著相关性。这说明生长性状和叶形性状没有直接的相关性,也说明所选的9个生长性状和叶形性状存在性状类型的冗余。将生长性状和叶形性状分成两类进行典型相关性分析发现两类性状的显著性均未达到显著或极显著水平,不符合开展典型相关性分析的试验要求。在今后的育种程序中,需要根据育种目标选择相关的性状开展综合选择,或通过主成分分析的方法对多种类性状进行降维,用更少的成分去代表更多类型的性状,提高育种选择效率[30]。通过性状间的相关性分析,可以清晰地了解性状间的关联,为优良家系的筛选提供理论基础[31]。
林以种为本,种以质为先。林木种质资源是林业种业发展的基础,了解种质资源的差异并从中挖掘优良种质是育种工作的核心内容,对种质的引种栽培与新种质创制等方面具有重要意义。优良家系的筛选方法根据树种的不同存在显著的差异[32],例如黄秀[33]主要利用方差分析模型,结合遗传增益和变异系数估算的方法开展5个种源樟树测定林的筛选分析;邹智滢[34]则更倾向于性状本身的差异分析,通过方差分析、主成分分析和聚类分析,筛选出综合指标最高的6个香樟Camphora officinarum家系,与本研究的方法相近。本研究采用TOPSIS法,通过赋予树高、胸径和冠幅表型相应的权重值,综合筛选出8个红豆树优良家系,为红豆树的良种选育提供材料。
植物表型性状变异是由遗传因素和环境因素共同作用的结果,在相同的生长环境、栽培管理条件下探究红豆树各家系和各来源不同性状的多样性才能更好地研究其遗传本质。本研究的不足之处在于,所涉及到的试验材料以省份作为产地,且其中三个省份在地理位置上是毗邻关系,难以准确说明不同来源的红豆树表型变异规律。考虑到某些产地可能处于省份交界处或地理距离较近,在今后的研究中,应在严格意义上的产地收集材料而不是广义的以省份替代,仅以省份作为种质资源的划分类别不能清晰地区分遗传差异对表型性状的影响;另一方面可以进行基于分子标记的遗传多样性分析,从遗传角度筛选红豆树优株与核心种质,为红豆树育种资源的挖掘奠定基础。
3.3 结论
红豆树的生长性状与叶形性状在各产地和家系间均存在丰富的差异,其中四川产地的红豆树特征明显,长势较差但叶片生长较好。表型性状间的相关性表明生长和叶形两类性状内表型性状两两之间均存在极显著正相关P< 0.01),但两类表型性状之间相关性不强。聚类分析中将38个红豆树家系聚成了5大类,其中福建产地的红豆树家系差异相对较大,聚类结果与地理位置相吻合。基于树高、胸径和冠幅等表型性状综合筛选得到8个优良红豆树家系。
-
![]()
图 1 基于9种表型的红豆树不同家系表型树状聚类图
SC-四川;FJ-福建;ZJ-浙江;JX-江西。
Figure 1. Cluster analysis of different populations of O. hosiei based on 9 phenotypic traits
表 1 红豆树采种产地与种质来源
Table 1 Originsites and seed groups of O. hosiei
编号 来源地 采集地点 编号 来源地 采集地点 JX1 江西省 抚州1号马头山镇 FJ1 福建省 南平市茫荡山自然保护区 JX2 江西省 抚州2号马头山镇 FJ2 福建省 南平市浦城县石陂镇 JX3 江西省 抚州zx6马头山镇 FJ3 福建省 南平市浦城县临江镇 JX4 江西省 抚州4号马头山镇 FJ4 福建省 宁德市柘荣县富溪镇 JX5 江西省 抚州尖峰1号广昌尖峰乡 FJ5 福建省 宁德市柘荣县宅中乡 ZJ1 浙江省 龙泉市锦溪镇 FJ6 福建省 宁德市柘荣县东源乡 ZJ2 浙江省 龙泉市八都镇 FJ7 福建省 宁德市柘荣县富溪镇 ZJ3 浙江省 龙泉市林业科学研究院 FJ8 福建省 宁德市柘荣县宅中乡 ZJ4 浙江省 龙泉市林业科学研究院 FJ9 福建省 宁德市福安市下白石镇 ZJ5 浙江省 龙泉市林业科学研究院 FJ10 福建省 宁德市福安市下白石镇 ZJ6 浙江省 龙泉市林业科学研究院 FJ11 福建省 宁德市柘荣县富溪镇 ZJ7 浙江省 龙泉市外窟 FJ12 福建省 宁德市柘荣县富溪镇 ZJ8 浙江省 龙泉市林业局 FJ13 福建省 宁德市柘荣县富溪镇 ZJ9 浙江省 龙泉市杨梅尖 FJ14 福建省 宁德市柘荣县富溪镇 ZJ10 浙江省 龙泉市八都镇 FJ15 福建省 宁德市柘荣县富溪镇 ZJ11 浙江省 龙泉市剑池街道 SC1 四川省 资阳市安岳县李家镇 ZJ12 浙江省 龙泉市江滨路 SC2 四川省 内讧市太平镇 ZJ13 浙江省 丽水市云和县西弄村 SC3 四川省 内讧市太平镇 ZJ14 浙江省 丽水市庆元县黄田镇 ZJ15 浙江省 丽水市庆元县黄田镇  下载: 导出CSV
下载: 导出CSV
表 2 不同产地红豆树生长性状和叶形性状的测定
Table 2 Determination of growth traits and leaf morphological traits in different O. hosiei populations
表型性状 江西 浙江 福建 四川 树高/m 3.36±0.09b 4.03±0.11a 4.12±0.05a 3.01±0.13b 胸径/cm 3.60±0.13b 4.16±0.07a 4.16±0.09a 3.07±0.15c 东西冠幅/m 1.76±0.06b 1.89±0.03ab 1.95±0.03a 1.31±0.07c 南北冠幅/m 1.72±0.06a 1.89±0.03a 1.91±0.03a 1.35±0.09b 分枝数量/个 7.51±0.26ab 8.20±0.16a 8.26±0.13a 7.06±0.30b 叶面积/cm2 15.55±0.39b 15.42±0.20b 16.78±0.25b 23.56±0.76a 叶周长/cm 13.33±0.18bc 13.09±0.11c 13.91±0.12b 16.21±0.30a 叶长/cm 9.65±0.09bc 9.37±0.05c 9.83±0.06b 11.57±0.19a 叶宽/cm 4.05±0.05c 4.19±0.03bc 4.28±0.03b 5.50±0.08a 注:数据列中不同小写字母表示在P<0.01水平存在极显著差异。
下载: 导出CSV
表 3 红豆树生长性状和叶形性状在产地间和家系间的方差分析
Table 3 Analysis of variance on growth and leaf morphological traits among O. hosiei populations and individuals
性状 产地间F值(Fp) 家系间F值(Fi) 树高 18.902** 2.774** 胸径 24.674** 11.881** 冠幅 0.864 1.096 分枝数 6.707** 3.712** 叶面积 52.184** 10.126** 叶周长 33.842** 7.118** 叶长 28.028** 13.881** 叶宽 106.173** 19.567** 注: **表示产地间和家系间均存在极显著差异(P<0.01)。
下载: 导出CSV
表 4 红豆树4个产地生长性状与叶形性状变异系数
Table 4 Coefficient of variation of growth and leaf morphological traits among O. hosiei populations
来源地 树高/% 胸径/% 东西冠幅/% 南北冠幅/% 分枝数/% 叶面积/% 叶周长/% 叶长/% 叶宽/% 江西 28.51 37.85 34.85 34.15 36.03 30.57 16.86 11.67 15.22 浙江 26.80 33.14 33.47 33.72 35.59 27.62 17.59 11.63 13.67 福建 22.94 38.90 27.64 25.93 30.64 31.24 18.31 13.41 15.65 四川 36.02 41.43 47.82 56.90 35.94 30.78 17.28 15.68 13.95
下载: 导出CSV
表 5 红豆树表型性状产地间和产地内的方差分量与表型分化系数
Table 5 Variance portion and phenotypic differentiation coefficient (VST) among population
性状 方差分量 方差分量百分比/% 表型分化系数(VST)/% 产地间(σ2t/s) 产地内(σ2s) 产地间(σ2t/s) 产地内(σ2s) 树高 0.24 0.06 19.22 5.04 79.21 胸径 0.07 0.68 2.96 29.59 9.10 东西冠幅 0.07 0.02 16.86 5.62 75.00 南北冠幅 0.06 0.02 13.79 5.67 70.89 分枝数 0.10 0.73 1.38 9.73 12.41 叶面积 13.68 2.56 35.02 6.56 84.23 叶周长 1.79 0.48 22.96 6.17 78.81 叶长 0.88 0.27 35.30 10.82 76.55 叶宽 0.41 0.08 49.88 9.98 83.33
下载: 导出CSV
表 6 红豆树表型性状相关性分析
Table 6 The correlation between traits in the O. hosiei populations
树高 胸径 东西冠幅 南北冠幅 分枝数 叶面积 叶周长 叶长 叶宽 树高 1.000 胸径 0.477** 1.000 东西冠幅 0.547** 0.681** 1.000 南北冠幅 0.491** 0.617** 0.783** 1.000 分枝数 0.171** 0.252** 0.317** 0.303** 1.000 叶面积 0.039 −0.042 0.019 0.013 0.014 1.000 叶周长 0.071* −0.001 0.026 0.016 0.028 0.879** 1.000 叶长 0.042 −0.004 0.028 0.030 0.006 0.667** 0.634** 1.000 叶宽 0.033 −0.078* 0.018 0.034 −0.025 0.724** 0.460** 0.622** 1.000 注:**表示极显著相关(P<0.01),*表示显著相关(P<0.05)。
下载: 导出CSV
表 7 8个优良家系的表型指标及排名
Table 7 Phenotypic indicators and rankings of 8 excellent families of O. hosiei
家系号 树高/m 胸径/cm 东西冠幅/m 南北冠幅/m 指标(CI) 排名 ZJ6 4.44±0.19a 5.23±0.29a 2.36±0.10a 2.38±0.11a 0.9564 1 FJ14 4.63±0.20a 5.26±0.24a 2.19±0.11a 2.09±0.10ab 0.9224 2 FJ12 4.56±0.18a 5.14±0.26a 2.13±0.10a 2.09±0.07ab 0.9080 3 ZJ5 4.42±0.16a 4.87±0.22a 2.21±0.10a 2.19±0.09ab 0.8853 4 ZJ8 4.23±0.19a 4.93±0.23a 2.13±0.10a 2.18±0.10ab 0.8629 5 FJ13 4.38±0.23a 4.97±0.32a 2.19±0.14a 1.95±0.14b 0.8621 6 FJ8 4.28±0.18a 4.99±0.28a 1.99±0.11a 2.02±0.10ab 0.8447 7 FJ6 4.09±0.13a 4.74±0.20a 1.94±0.08a 1.97±0.07ab 0.7841 8 入选均值 4.38 5.02 2.14 2.11 注:同一列中不同小写字母表示在P<0.01水平存在极显著差异。
下载: 导出CSV
-
[1] 李峰卿,周志春,楚秀丽,等. 红豆树优树子代遗传多样性及与生长相关性分析[J]. 林业科学研究,2018,31(1):42 − 50. [2] 陈存及,陈伙法. 阔叶树种栽培[M]. 北京:中国林业出版社,2000:132 − 134. [3] 楚秀丽,付艳茹,严巍. 珍稀植物红豆树资源保育及精细化培育研究进展[J]. 中国野生植物资源,2021,40(10):61 − 65. doi: 10.3969/j.issn.1006-9690.2021.10.010 [4] 王明彬,韦小丽,韦忆,等. 川黔地区濒危植物红豆树种群结构与数量动态特征[J]. 广西植物,2024,44(1):179 − 192. [5] 蔡勇. 鄂西红豆树育苗技术初步研究[J]. 林业勘察设计,2007(2):97 − 100. doi: 10.3969/j.issn.1004-2180.2007.02.028 [6] ZHANG R,ZHOU Z,DU K. Genetic diversity of natural populations of endangered Ormosia hosiei,endemic to China[J]. Biochem Syst Ecol,2012,40:13 − 18. doi: 10.1016/j.bse.2011.09.005
[7] 尤根彪,叶和军,焦洁洁,等. 浙江丽水白云山红豆树人工林的径级结构和空间分布格局[J]. 广西植物,2017,37(6):799 − 805. doi: 10.11931/guihaia.gxzw201609030 [8] 李峰卿,周志春,谢耀坚. 3个小流域红豆树天然居群的遗传多样性和遗传分化[J]. 分子植物育种,2017,15(10):4263 − 4274. [9] KAUSHIK N,DESWAL R P S,MALIK S,et al. Genetic variation and heritability estimation in Jatropha curcas L. progenies for seed yield and vegetative traits[J]. Journal of Applied and Natural Science,2015,7(2):567 − 573. doi: 10.31018/jans.v7i2.646
[10] PINYOPUSARERK K,WILLIAMS E R. Range-wide provenance variation in growth and morphological characteristics of Casuarina equisetifolia grown in Northern Australia[J]. Forest Ecology and Management,2000,134(1-3):219 − 232. doi: 10.1016/S0378-1127(99)00260-1
[11] VIZCAINO-PALOMAR N,IBANEZ I,BENITO-GARZON M,et al. Climate and population origin shape pine tree height-diameter allometry[J]. New Forests,2017,48:363 − 379. doi: 10.1007/s11056-016-9562-4
[12] 贾庆彬,刘庚,赵佳丽,等. 红松半同胞家系生长性状变异分析与优良家系选择[J]. 南京林业大学学报(自然科学版),2022,46(4):109 − 116. [13] 王云鹏,张蕊,周志春,等. 10年生木荷生长和材性性状家系变异及选择[J]. 南京林业大学学报(自然科学版),2020,44(5):85 − 92. [14] 潘艳艳,梁德洋,郭婧,等. 日本落叶松不同种源及家系生长性状变异分析[J]. 北京林业大学学报,2018,40(11):19 − 27. [15] 陈清根. 不同种源红豆树遗传变异分析及选择[J]. 林业勘察设计,2021(3):19 − 21,25. [16] 崔珺,孙梦,程瀚远,等. 不同地理种源红豆树幼苗叶片功能性状与碳氮磷化学计量分析[J]. 凯里学院学报,2023,41(3):40 − 50. doi: 10.3969/j.issn.1673-9329.2023.03.007 [17] 李颖,张树航,郭燕,等. 中国板栗9个结果母枝相关表型性状遗传多样性研究[J]. 园艺学报,2019,46(3):453 − 463. [18] 李峰卿,陈焕伟,周志春,等. 红豆树优树种子和幼苗性状的变异分析及优良家系的初选[J]. 植物资源与环境学报,2018,27(2):57 − 65. [19] 葛颂,王明庥,陈岳武. 用同工酶研究马尾松群体的遗传结构[J]. 林业科学,1988,24(4):399 − 409. [20] 李斌,顾万春,卢宝明. 白皮松天然群体种实性状表型多样性研究[J]. 生物多样性,2002,10(2):181 − 188. doi: 10.3321/j.issn:1005-0094.2002.02.008 [21] 王艺林,吕东,刘贤德,等. 5个种源文冠果果实和种子表型性状变异分析[J]. 经济林研究,2018,36(3):38 − 47. [22] 穆怀志,王清玉,张玥,等. 基于叶片和果实性状的糠椴天然居群表型变异及多样性分析[J]. 植物研究,2023,43(06):826 − 834. [23] 程琳,肖建军,覃孟哲,等. 杉木木材品质分析及优良种质筛选[J]. 福建林业科技,2023,50(3):9 − 13,21. [24] BOGDAN S,KATICIC-TRUPCEVIC I,KAJBA D. Genetic variation in growth traits in a Quercus robur L. open-pollinated progeny test of the Slavonian provenance[J]. Silvae Genetica,2004,53(1-6):198 − 201. doi: 10.1515/sg-2004-0036
[25] 周连第,兰彦平,韩振海. 板栗品种资源分子水平遗传多样性研究[J]. 华北农学报,2006,21(3):81 − 85. doi: 10.3321/j.issn:1000-7091.2006.03.019 [26] WILLS C. Principles of population genetics[M]. Sunderland:Sinauer Associates,Inc. Publishers,2007:382.
[27] 王娜,杨会娜,肖健. 蒙古栎主要种群表型性状变异分析及多样性研究[J]. 林业与生态科学,2023,38(3):269 − 274. [28] 刘亚斌,郭素娟,孙传昊. 基于巢式分组设计的板栗刺苞与坚果形态多样性分析[J]. 中南林业科技大学学报,2020,40(10):51 − 60. [29] FUKATSU E,HIRAOKA Y,MATSUNAGA K,et al. Genetic relationship between wood properties and growth traits in Larix kaempferi obtained from a diallel mating test[J]. Journal of Wood Science,2015,61:10 − 18. doi: 10.1007/s10086-014-1436-9
[30] 段红静. 杉木种质资源遗传多样性评价及重要性状的全基因组关联分析[D]. 北京:北京林业大学,2017. [31] 罗芊芊,楚秀丽,李峰卿,等. 5年生南方红豆杉生长和分枝性状家系变异与选择[J]. 林业科学研究,2020,33(1):136 − 143. [32] 杨孟晴. 两种青冈家系遗传变异分析及优良家系选择[D]. 合肥:安徽农业大学,2023. [33] 黄秀美. 樟树优树子代生长和形质性状综合选择[J]. 林业勘察设计,2021,41(1):25 − 29. [34] 邹智滢. 香樟不同家系生长特性与初步选择的研究[D]. 福州:福建农林大学,2016.
计量
- 文章访问数: 11
- HTML全文浏览量: 1
- PDF下载量: 0
